Rollen til motorproteiner i celledeling
Av Jonathan McLatchie, 8. mai 2024, 2:22. Oversatt herfra
Bilde 1. Celledeling
Et av de mest forbløffende eksemplene på molekylære maskiner i cellen er den mitotiske spindelen som styrer segregeringen av kromosomer under celledelingen. Du kan se den forbløffende prosessen med mitotisk celledeling utfolde seg i denne animasjonen:
Et avgjørende aspekt ved mitose er bevegelse av kromosomer - opprinnelig oppstillingen av sammenkoblede søsterkromatider under prometafase og metafase, og deretter separasjonen av disse søsterkromatidene under anafase. Men hvordan utøver spindelen kraft på kromosomene? Og hvordan festes kromosomene til spindelen slik at de kan flyttes til riktig sted til riktig tid? Her vil jeg gi en introduksjon til denne forbløffende prosessen.
Mikrotubuler
To strukturelle komponenter utgjør den mitotiske spindelen - to store strukturer, funnet ved polene, kalt sentrosomer, og proteinfilamenter kjent som mikrotubuli. Fra hver av sentrosomene stråler spindelmikrotubuli, som binder seg til et kromosom (kinetokor mikrotubul), eller et mikrotubul som stammer fra det andre sentrosomet (polar mikrotubul), eller cellens plasmamembran (astral mikrotubul).
Underenheten til mikrotubuli er heterodimere av tubulin, bestående av a-tubulin og ß-tubulin. Disse heterodimerene passer sammen for å lage lineære protofilamenter, hvorav 13 er justert side om side for å danne en mikrotubul. På grunn av asymmetrien til tubulin heterodimerer, har mikrotubulen egenpolaritet. Plussenden (som strekker seg mot kromosomene) består av ß-tubulin-underenheter, mens minusenden (som er forankret ved sentrosomet) består av a-tubulin-underenheter.
I de tidlige mitotiske stadiene samles mikrotubul og demonterer raskt (mediert av tilsetning og fjerning av tubulin heterodimere), et fenomen kjent som dynamisk ustabilitet (prosessene for depolymerisering og polymerisering blir henholdsvis referert til som katastrofe og redning). Ved denne prosessen undersøker mikrotubuli tilfeldig cellen, til de ved en tilfeldighet møter målet. Reguleringen av mikrotubul-katastrofe og -redning er knyttet til hydrolyse av GuanosinTriFosfat (GTF, et nukleotid som er bundet til tubulin i mikrotubulens gitter. Hver tubulin-underenhet er i stand til å binde seg til ett molekyl GTF. Under polymerisasjon tilsettes tubulin -underenheter bundet til GTF, til den voksende enden av mikrotubulen, og danner en GTF-hette som stabiliserer mikrotubulen og fremmer vekst.
 Når mikrotubulen fortsetter å polymerisere, gjennomgår GTF-bundne tubulin-underenheter hydrolyse, og konverterer GTF til Guanosin-Difosfat (GDF). Når de er hydrolysert, blir tubulin -underenhetene mindre stabile, og mikrotubulen er mer utsatt for depolymerisering. Når du når en kritisk konsentrasjon av GDF-bundet tubulin i mikrotubulienden, oppstår ‘katastrofe’, noe som fører til rask depolymerisering. Mikrotubulusredning involverer utveksling av GDF-bundet tubulin for GTP-bundet tubulin, og fremmer dermed mikrotubulusvekst. Enkelte cellulære faktorer, så som Mikrotubul-Assosierte Proteiner (MAP) og motoriske proteiner, kan lette denne prosessen ved å fremme inkorporering av GTP-bundet tubulin eller ved å beskytte GTP-hetten mot hydrolyse. For eksempel er kinesin-8-proteiner pluss-mål-rettede motoriske proteiner som destabiliserer mikrotubuler. (1) Et annet motorisk protein, kinesin-13, er toveis (kan bevege seg i retning av enten pluss eller minus ender av mikrotubulen), i kontrast til de fleste motorer som er ensrettet. De er derved i stand til å fremme depolymerisering av en mikrotubul fra begge sider. (2) Aktiviteten til kinesin-13-proteiner er "regulert av distinkt målretting av regioner i spindelen, ved regulatoriske fosforyleringshendelser og av interaksjoner med forskjellige bindingspartnere." Disse bemerkelsesverdige kinesin-13 motorene binder enden av mikrotubulen og utløser en konformasjonsendring som resulterer i mikrotubul depolymerisering. (4)
Når mikrotubulen fortsetter å polymerisere, gjennomgår GTF-bundne tubulin-underenheter hydrolyse, og konverterer GTF til Guanosin-Difosfat (GDF). Når de er hydrolysert, blir tubulin -underenhetene mindre stabile, og mikrotubulen er mer utsatt for depolymerisering. Når du når en kritisk konsentrasjon av GDF-bundet tubulin i mikrotubulienden, oppstår ‘katastrofe’, noe som fører til rask depolymerisering. Mikrotubulusredning involverer utveksling av GDF-bundet tubulin for GTP-bundet tubulin, og fremmer dermed mikrotubulusvekst. Enkelte cellulære faktorer, så som Mikrotubul-Assosierte Proteiner (MAP) og motoriske proteiner, kan lette denne prosessen ved å fremme inkorporering av GTP-bundet tubulin eller ved å beskytte GTP-hetten mot hydrolyse. For eksempel er kinesin-8-proteiner pluss-mål-rettede motoriske proteiner som destabiliserer mikrotubuler. (1) Et annet motorisk protein, kinesin-13, er toveis (kan bevege seg i retning av enten pluss eller minus ender av mikrotubulen), i kontrast til de fleste motorer som er ensrettet. De er derved i stand til å fremme depolymerisering av en mikrotubul fra begge sider. (2) Aktiviteten til kinesin-13-proteiner er "regulert av distinkt målretting av regioner i spindelen, ved regulatoriske fosforyleringshendelser og av interaksjoner med forskjellige bindingspartnere." Disse bemerkelsesverdige kinesin-13 motorene binder enden av mikrotubulen og utløser en konformasjonsendring som resulterer i mikrotubul depolymerisering. (4)
Bilde 2. Ulike faser under mitose (celledeling)
Motorproteiner
I 2014 publiserte Discovery Institute en animasjon av Molecular Motor Kinesin, som bærer cellulær last langs mikrotubul-spor og drevet av ATP -hydrolyse:
Cellens arbeidshest: Kinesin -lenke
Kinesin beveger seg typisk i retning av plussenden av mikrotubulen, mot periferien av cellen. En annen form for motorprotein, kalt dynein, beveger seg generelt mot minusenden av mikrotubulen, dvs. mot cellesenteret. Mens kinesin-motoriske proteiner går trinn for trinn ved å plassere den ene foten foran den andre, beveger dynein seg med en svingende brokryssende-mekanisme.
En av de mest utrolige funksjonene til motoriske proteiner i cellen er deres rolle i monteringen og funksjonen til den mitotiske spindelen under eukaryotisk celledeling.(5) Se for deg en robotfabrikk som samler og organiserer cellen som forberedelse til å gjennomgå deling, og bidra til å lette den kontrollerte segregering av det genetiske materialet inne i dattercellene. Dette er representert i den (svært forenklede) figuren nedenfor:
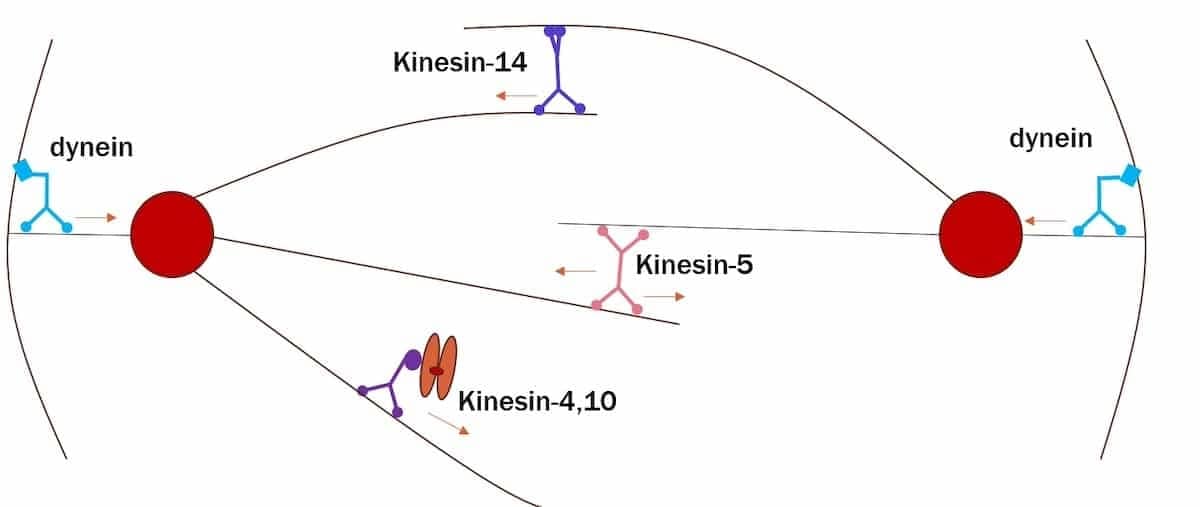 Bilde 3. Kinesin variamter-wikepedia
Bilde 3. Kinesin variamter-wikepedia
Som vist på figuren, kan de motoriske proteinene som er involvert i organiseringen av den mitotiske spindelen deles inn i fire hovedklasser: kinesin-5, kinesin-14, kinesins-4 og 10 og dynein. Kinesin-5 er rettet mot plussenden og glir fra hverandre mikrotubuli som har sine polariteter orientert i motsatte retninger (dvs. antiparallelle mikrotubuli). Dermed bidrar kinesin-5 til spindelforlengelse og bipolaritet ved å skyve spindelpolene fra hverandre. (6) På den annen side er kinesin-14 motorer rettet mot minus-enden av mikrotubulen. De har hver et motorisk domene i tillegg til andre domener som forbinder seg med et annet mikrotubul. (7) De krysslinker antiparallelle interpolare mikrotubuler ved spindelens midtsone, og trekker derved polene mot hverandre.
En annen gruppe kinesiner, kinesin-4 og kinesin-10 (samlet kjent som kromokineriner), spiller en kritisk rolle i posisjonering og segregering av kromosomer under mitose.(8) Jobben til kromokinesiner er å flytte kromosomene til sine rette posisjoner slik at hver dattercelle vil motta et riktig antall kromosomer.
Den siste gruppen av motoriske proteiner involvert i mitose er dynein, som har sine motoriske domener assosiert med mikrotubuli, som stammer fra sentrosomene, og deres lastbindende domener bundet til proteiner innebygd i cellebarken. Bevegelsen av dynein utøver en trekkraft på mikrotubulen minus ender og trekker sentrosomene i retning av cellebarken.(9) Denne bevegelsen sikrer at hver dattercelle får et komplett sett med kromosomer. Legg merke til at selv om jeg for enkelhets skyld bare har vist en mikrotubul som stammer fra hvert centrosom mot cellebarken, ville det i det virkelige liv være mange flere enn dette.
Utrolig prosjektering
I et hvilket som helst annet erfaringsområde ville vi, når vi observerer en robotfabrikk som denne som opererer i en nanoskala, umiddelbart tilskrive den til design. Ingeniørfaget for den eukaryote celledelingssyklusen er forbløffende på alle nivåer. Dessuten er disse molekylære motorene absolutt uunnværlige for vellykket celledeling i eukaryoter og utgjør dermed et ikke-reduserbart komplekst system. Det peker derfor kraftig og utvetydig på bevisst intensjon.
Oversettelse, via google oversetter, og bilder ved Asbjørn E. Lund
---